Bases biológicas de la agresividad
Bases
biológicas de la conducta agresiva
Formas
y tipos fundamentales de violencia y agresión
Somos conscientes de que los
actos de violencia y agresión son muy variados y su tipología puede
establecerse siguiendo varios criterios clasificatorios (Brain, Olivier, Mos,
Benton, y Bronstein, 1998),
A) Según los
modos de la agresión
Violencia Directa-Violencia
Indirecta: Si se tiene en cuenta el modo de producirse la agresión puede
hablarse de violencia estructural o indirecta y violencia directa o personal.
En la violencia directa
(personal) los actos destructivos son realizados por personas o colectivos
concretos y se dirigen también a personas o grupos igualmente definidos.
En la violencia indirecta o
estructural no hay actores concretos de la agresión; en este caso la
destrucción brota de la propia organización del grupo social sin que tenga que
haber necesariamente un ejecutor concreto de la misma.
B)
Según sus actores
Si en vez de atender a la
forma de producirse la violencia, nos fijamos en los actores de la agresión y
en los sujetos de la sufren entonces encontramos los siguientes actos
violentos:
- De un individuo contra sí
mismo (suicidio).
- De un individuo contra otro
individuo (crimen pasional).
- De un individuo contra un
grupo (delitos contra la sociedad).
- De un grupo contra un
individuo (la pena de muerte).
- De un grupo contra otro
grupo (la guerra, el terrorismo)
C)
Según otros criterios
Aparte de los dos criterios
clasificatorios precedentes existen otros que permiten matizar y completar la
tipología de la violencia. Así, frente a la violencia espontánea de un
individuo o de una masa, está la violencia organizada de las guerras.
Por último, teniendo en cuenta el «mecanismo » desencadenante puede hablarse de una violencia normal y una violencia patológica, esta última puede ser provocada por alteraciones psíquicas primarias o por modificaciones anormales del funcionamiento cerebral.
Como mencionamos al comienzo
de este apartado existen otras clasificaciones de destacada importancia, como
la propuesta por Moyer (1976):
1. Agresión predatoria
(conductas de ataque motivadas).
2. Agresión inter-machos
(violencia física o conducta de sumisión exhibida por los machos mutuamente).
3. Agresión inducida por el
miedo (respuestas biológicamente programadas de modo que se actúa de forma
agresiva hacia cualquier clase de confinamiento forzado).
4. Agresión territorial
(conducta de amenaza o ataque que se muestra hacia una invasión del territorio
propio, o conducta de sumisión y retirada tras enfrentarse con el intruso).
5. Agresión maternal
(conducta agresiva mostrada por las hembras cuando un intruso se acerca a sus
crías).
6. Agresión irritable
(agresión e ira dirigidas hacia un objeto cuando el agresor se siente frustrado,
herido, deprivado o estresado).
7. Agresión relacionada con
el sexo (elicitada por los mismos estímulos que disparan la respuesta sexual).
8. Agresión instrumental (la
que conduce al individuo a obtener una recompensa mediante el acto agresivo). 
Modelo neurobiológico:
Evidencias
de alteraciones prefrontales:
Tras el accidente ocurrido a
Phineas Gage en 1848 se ha avanzado significativamente en el conocimiento neuropsicológico
y neurofisiológico de la agresión; el radical cambio comportamental luego del
accidente pudo explicarse -tras la muerte de Gage- por las evidentes lesiones
en la áreas anteriores del cerebro.
Éste primer caso sumado con un
número significativo de evidencias empleando técnicas de Neuroimágen
estructurales y funcionales como la Resonancia Magnética (RM) o la Tomografía
por Emisión de Positrones (TEP) han corroborado el vínculo entre córtex
prefrontal (CPF) y agresión, comprobando que las alteraciones en esta región cerebral
se asocian con fallas de regulación y control emocional, impulsividad irritabilidad,
fallas en la planeación de la conducta y del comportamiento ético y moral
(Miller & Cummings, 2007).
Adicionalmente, el empleo de técnicas
de Neuroimágen ha permitido evidenciar los circuitos que se establecen entre el
CPF y estructuras subcorticales como la amígdala, el hipocampo o el hipotálamo,
lo que demuestra la importancia de esta región cortical en la regulación cognitivo-emocional
(Alcazar, et al. 2012; Pinel, 2007; Portellano, 2005).
Un adecuado control emocional favorece la inhibición de la respuesta agresiva, lo cual está directamente asociado con la integridad del CPF y las Funciones Ejecutivas (FE), las cuales están asociadas con las capacidades autoevaluación, inhibición, flexibilidad y solución de problemas (Navas & Muñoz, 2004; Portellano, 2005; Alcázar, Verdejo, Bouso & Bezos, 2010).
Las funciones de planeación y
organización temporal del comportamiento se asocian con el córtex dorsolateral;
al evidenciarse apatía, desinterés o fallas atencionales, se asocia con la
región ventromedial, mientras la desinhibición, alta reactividad emocional e inestabilidad
afectiva emocional se asocian con la región orbitofrontal (Miller, Cummings,
2007; Arnedo et al. 2012).
Una dificultad que surge al
interpretar al CPF como sustrato importante en el control de la agresión ha
sido que la amplitud de la lesión en estas regiones corticales podría
acompañarse de un funcionamiento atípico de estructuras subcorticales, afectar
las interconexiones y el equilibrio relativo de la actividad
cortico-subcortical, lo cual complejiza el análisis del comportamiento cerebral
del sujeto agresivo. (Alcázar et al. 2010).

Neurociencia
de la agresión: Alteraciones a nivel subcortical
Circuitos que se establecen
entre estructuras límbicas como la amígdala, el hipocampo o el hipotálamo, y el
CPF, regulan y controlan la expresión emocional. Se ha demostrado que alteraciones
subcorticales alteran el condicionamiento de miedo y otras formas de aprendizaje
relacional dependiente de estructuras subcorticales, sugiriendo alteraciones
funcionales en las conexiones subcórtico-corticales que participan en el
control emocional, las cuales son deficiencias que caracterizan a los
delincuentes agresivos reactivos (Alcázar et al. 2010).
Las alteraciones funcionales
subcorticales sugieren que la agresión reactiva podría estar relacionada con la
falta de inhibición prefrontal, que a su vez podría relacionarse con una
disminución en la actividad serotoninérgica del circuito límbicoorbitofrontal.
Adicionalmente, se postula
que la región insular desempeña un papel importante en el reconocimiento de las
emociones propias y de otros, lo cual está relacionado con la empatía (Moya, Herrero
& Bernal, 2010).
Desde esta perspectiva, la interacción
social es esencial para el ser humano, y el desarrollo de habilidades como la
empatía entendida como la capacidad para sentir el dolor del otro estaría
relacionada con la acción gradual de un conjunto de circuitos neuronales que le
permitan al individuo establecer la experiencia emocional de otro individuo como
propia; al presentarse esta situación, neuronas en espejo ubicadas en regiones límbicas,
el cíngulo anterior, el córtex premotor, el giro frontal inferior y regiones
prefrontales comienzan a disparar momentos antes de la experiencia emocional,
hipotetizándose que alteraciones funcionales en esta red neuronal estaría presente
en individuos agresivos y en aquellos que no pueden detener la conducta
agresiva luego que se ha iniciado (Oberman, Pineda & Ramachandran, 2006)
Finalmente, un estudio que permite integrar los datos anteriormente expuestos fue realizado por Cornet, De Kogel, Ninkam, Raine y Van der Laan (2013), en el cual llevaron a cabo una serie de análisis comparativos con sujetos en los cuales predominaban respuestas de agresión afectiva (impulsiva) o agresión premeditada (depredadores). Empleando imágenes de Resonancia Magnética Funcional (RMf) lograron observar en agresores afectivos una disminución en la respuesta de la glucosa a nivel orbitofrontal, una reducción significativa en la sustancia gris del CPF, mayor activación amigdalina asociada con un volumen reducido de esta misma estructura y una hipoactividad observada en el flujo sanguíneo cerebral regional (FSCr) en la formación hipocampal, el giro hipocampal y el estriado –así como en la amígdala y el CPF- ante la presentación de estímulos afectivos; estos datos son consistentes con las evidencias conductuales de mayor impulsividad, reactividad emocional y dificultades para el reconocimiento de expresiones emocionales de miedo (Raine, Buchsbaum, Stanley, Lottenberg, Abel & Stodard, 1994; Raine, Stoddard, Bihrle & Buchsbaum, 1998; Raine, Lencz, Bihrle, LaCasse & Colleti, 2000).
Neurotransmisores
involucrados con la agresividad
A pesar que se ha evidenciado
que lesiones tanto en estructuras corticales como subcorticales pueden favorecer
la agresión, en ocasiones las alteraciones tienen un carácter más molecular que
estructural. En los siguientes apartados se expondrán las principales
sustancias neurotransmisoras, receptores y hormonas relacionadas con la
agresión
Serotonina
(5hidroxitriptamina)
La serotonina es un
neurotransmisor evolutivamente mantenido desde los peces al hombre, donde se la
encuentra en varias partes del organismo como la sangre o el sistema digestivo.
En el cerebro, la serotonina
forma un sistema muy distribuido con neuronas cuyos axones se originan en el
rafe y se distribuyen luego en áreas claves de la corteza límbica (hipocampo,
amígdala, hipotálamo) o sensorial- cognitiva (frontal, prefrontal, temporal, parietal)
La
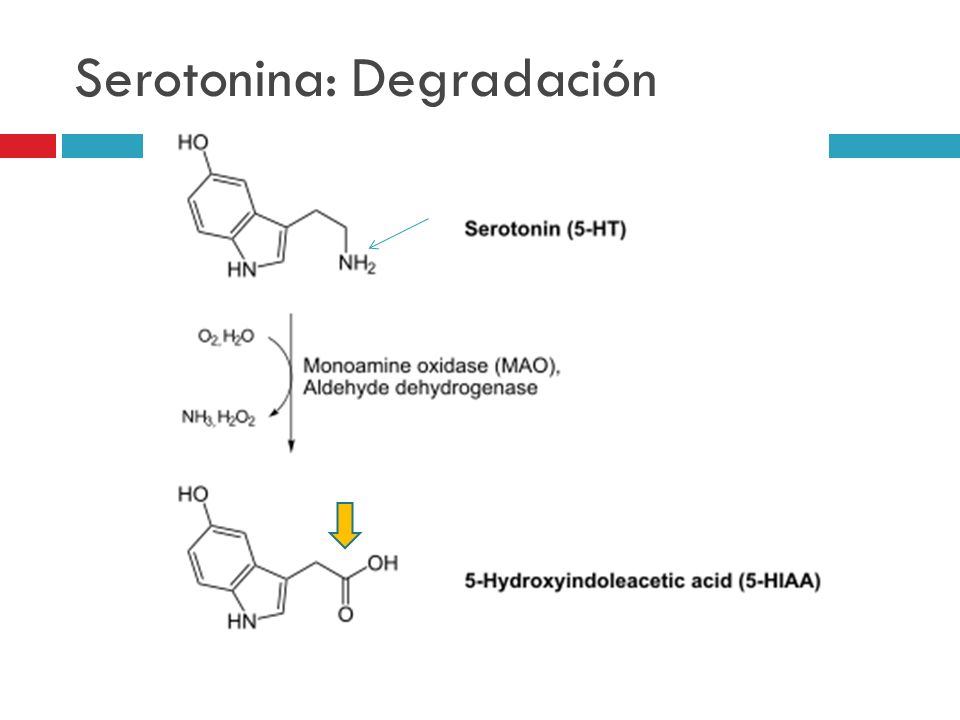
enzima que comienza la síntesis de serotonina es la triptofano hidroxilasa
(TPH) que representa el paso limitante. La serotonina liberada de las vesículas
en la sinapsis es recaptada por el recaptador de serotonina (5HTR) y en parte
metabolizada por la monoaminooxidasa, sobre todo la forma A (MAO A), que da
como resultado el metabolito 5 hidroxi indol acético (5 HIAA). A su vez, en el espacio
postsináptico la serotonina actúa sobre receptores de los cuales se ha
identificado ya 14 tipos diferentes (5HT 1 a 7, con subtipos A, B, C, o 1, 2,
etc., en algunos casos).
Estos receptores, excepto el
5HT5, activan segundos mensajeros como la adenil ciclasa, que a través de otras
kinasas fosforilan señales intracelulares que finalmente se traslocan al núcleo
y activan genes que aumentan la expresión de enzimas o factores de crecimiento,
generando cambios plásticos cerebrales.
Como se mencionó
anteriormente, la 5- hidroxitriptamina o serotonina (5-HT) es el
neurotransmisor más relacionado con la agresión; el principal precursor de 5-HT
es el triptófano, el cual se encuentra en alimentos como las carnes, verduras o
cereales (Purves, 2004).
Se ha descubierto, tanto en
modelos animales como en humanos, que dietas ricas en triptófano disminuyen la
ocurrencia de acciones agresivas favoreciendo sensaciones de bienestar, mientras
que la alimentación escasa en triptófano aumenta la irritabilidad y la agresión
en individuos altamente agresivos (Moya, 2010).
Adicionalmente, disminuciones
en los niveles de este neurotransmisor se asocian con aumentos en las reacciones
agresivas, impulsividad y disminución de la evaluación de riesgo.
La relación que se establece entre
menores niveles de 5-HT y mayor probabilidad de presentar conductas agresivas
ha sido evidenciada al realizar análisis en líquido cefalorraquídeo (LCR) en
individuos agresivos, encontrando menores concentraciones de metabolito para
5-HT, el ácido 5-hidroxidolacético (5-HIAA), lo que de acuerdo con las
investigaciones sugiere que la actividad serotoninérgica estaría disminuida en
regiones cercanas a los ventrículos cerebrales y el córte prefrontal (Miczek,
De Almeida, Kravitz, Rissman, De Boer & Raine, 2007).

Las
proteínas intermediarias del metabolismo de la serotonina y las conductas
agresivas
Sobre principios de la década de los
ochenta, el grupo de investigación de M. Asberg en el Instituto Karolinska, en
Estocolmo, estudiando la psicobiología del suicidio, separó un grupo de pacientes
con intento violento de autoeliminación (IAE), observando que estos tenían
menores concentraciones de 5-HIAA en líquido cefalorraquídeo, proporcionando
una fuerte evidencia para la hipótesis de que las conductas violentas cursan
con un metabolismo reducido de serotonina.
Se demostró luego que individuos
encarcelados por agresiones violentas, por ejemplo, mostraban el mismo cambio
metabólico.
Inversamente, estrategias farmacológicas
experimentales que aumentan los niveles de serotonina con el uso de
precursores, la inhibición del recaptador o el uso de agonistas de receptores
5HT1A y 5HT1B, han logrado reducir la agresividad en roedores
Se ha demostrado experimentalmente que
la genética de los fenotipos agresivos puede ser mediada por alteraciones de
los genes que codifican para la enzima de la síntesis de serotonina, la 5HTP,
la enzima de degradación, la MAO A o por el agonismo de un tipo de receptores,
los 5HT1A
El caso de la
MAO A
A comienzos de la década de los noventa
se describió una familia holandesa con una mutación en el gen para MAO A cuyos
miembros masculinos mostraban un patrón de comportamiento criminal, violento e
impulsivo, por generaciones.
El gen de MAO A tiene un alto
polimorfismo que influye marcadamente sobre la eficiencia transcripcional: la
enzima tiene una alta expresión en el caso de 3,5 ó 4 alelos (MAO A-H), siendo
su expresión baja en el caso de 2 ó 3 alelos (MAO A-L).
Dado que la MAO A es la enzima de
degradación de la serotonina, un aumento de su actividad (genotipo MAO A-H) se
traduce en menores niveles de serotonina, por lo que estos datos sobre el genotipo
de MAO A coinciden con los estudios que miden directamente los metabolitos del
neurotransmisor (5-HIAA).
Así mismo, agonistas de los receptores
5-HT1A, 5HT1B y 5 HT2A disminuyen las conductas agresivas en roedores mientras
que los antagonistas para estos mismos receptores aumentan la frecuencia de
éstas conductas, estableciéndose una relación positiva entre una mayor densidad
y sensibilidad de estos receptores, en especial 5-HT1A, en regiones corticales
y límbicas para los animales con mayor tendencia a la agresividad rasgo.
·
Aminoácidos y agresión(GABA Y
Glutamato)
Una de las primeras teorías
que se propuso para explicar la agresión se relacionó con el
desequilibrio neuroquímico
entre la actividad excitatoria e inhibitoria (Sustková, Vávrová & Krsiak,
2009). Así, se sugirió que un aumento en la actividad glutamatérgica o una
disminución en la actividad GABAérgica podrían facilitar la respuesta agresiva
(Monroe, 1978; Sustková, Vávrová & Krsiak, 2009).
Al respecto, en un estudio
realizado con ratas Wistar por Miczek, Fish, De Bold, (2007) se demostró que los
animales agresivos mostraban mayores niveles de glutamato y menores de GABA en
la corteza prefrontal, el hipotálamo y la amígdala en comparación con los no
agresivos, y cuando se administraban fármacos antagonistas de glutamato o agonistas
de GABA la respuesta agresiva disminuía.
Estudios con humanos han
demostrado que los fármacos empleados para el tratamiento de la epilepsia que
actúan principalmente sobre el sistema GABAérgico no sólo mejoran las crisis
convulsivas sino que también son útiles para disminuir la respuesta agresiva. conjunto
de estructuras que involucran regiones hipotalámicas anteriores, laterales y ventromediales,
el septum y la sustancia negra, son fuertemente influenciadas por los efectos excitatorios
e inhibitorios del glutamato y el GABA.
Estudios realizados con
roedores han demostrado que la administración de agonistas de los receptores glutamatérgicos
en estas regiones cerebrales provocan un aumento en el ataque ofensivo, evidenciándose
también una reducción de los niveles de GABA en estas regiones, así como en el hipocampo
y la amígdala (Moya, 2010).
Adicionalmente, se ha
descrito que las benzodiacepinas en algunas ocasiones funcionan como
facilitadores o inhibidores de la conducta agresiva (Miczek et al. 2007).
·
Catecolaminas(Dopamina ,
Noradrenalina, Adrenalina)
Pese a que la MAO participa
en el metabolismo de la 5-HT, también lo hace para la dopamina, noradrenalina y
adrenalina, las cuales comparten a la tirosina como precursor (Purves, 2004).
Así, la inhibición de la acción de estas enzimas conllevaría a un desequilibrio
neuroquímico de las aminas y el resultado de esta alteración podría ser una
mayor propensión del individuo a ejecutar conductas agresivas. 
Dopamina:
Respecto a lo anterior, se
sugiere que un aumento en los niveles de dopamina podría estar relacionado con
la intensificación de la agresión; esto ha podido evidenciarse en modelos
animales al inhibir la expresión para el transportador para dopamina (DAT) y
medir los niveles de neurotransmisor, encontrando que los individuos más
agresivos eran aquellos que mostraban mayores niveles de dopamina en el momento
de la agresión (Rodriguez, Chu, Caron & Wetsel, 2004).
Así mismo, se ha demostrado
que los receptores dopaminérgicos D2 tienen un papel importante en la conducta
agresiva, encontrándose que al ser estimulada la expresión del alelo
correspondiente a la forma corta del gen que codifica este receptor (D2) se
incrementan los niveles de agresión.

Así, se propone que la
administración de agonistas para receptores tanto D1 como D2 (como por ejemplo las
anfetaminas y la cocaína) inducen en modelos animales como en sujetos humanos
intranquilidad e incrementos en la irritabilidad, hostilidad y conducta
agresiva, observándose el efecto opuesto cuando se administran neurolépticos
típicos, como el haloperidol, los cuales son antagonistas para estos receptores,
en especial el D2 (De Almeida, Ferrari, Parmigiani & Miszek, 2005).
Noradrenalina:
Por su parte, se ha
relacionado al sistema noradrenérgico con el incremento de la actividad generalizada
del organismo, incluida la conducta agresiva, con efectos directos a nivel del
sistema nervioso central, autónomo y sistema endocrino; en estudios con modelos
animales se ha podido observar que aumentos en la actividad noradrenérgica disminuye
el umbral para iniciar el ataque agresivo y que los animales a los cuales se
les manipuló genéticamente para que no pudieran expresar las enzimas MAO y COMT
aumentaban los niveles de noradrenalina con el subsecuente aumento en la
expresión de conductas agresivas.
Finalmente, aquellos animales
incapaces de convertir dopamina en noradrenalina mostraron una disminución en
la agresión (Marino, Bourdelat, Cameron & Weinshenker, 2005).
Neuroendocrinología
de la Agresión
Testosterona:
Clásicamente se ha descrito
que los hombres tienen mayor tendencia a comportarse de formas agresivas en
comparación con las mujeres, estableciendo el argumento que una mayor liberación
de Testosterona en los machos los predispone a comportarse de forma agresiva.
Aunque, ciertamente, los
machos desde el periodo gestacional experimentan varios picos de liberación de
Testosterona que aumenta los niveles se esta hormona y favorece los procesos de
masculinización primaria y secundaria, no puede concluirse una relación causal
entre Testosterona y agresión; sin embargo, si es factible hipotetizar efectos
indirectos de un grupo de hormonas sobre la actividad sináptica, en las que se
incluye la Testosterona, la Vasopresina o la Oxitocina.
Los modelos animales han demostrado
que la administración exógena de Testosterona disminuye el umbral para el
inicio y aumenta los niveles de agresión, razón por la cual se ha sugerido que
los altos niveles de Testosterona facilitan la conducta agresiva.
Sin embargo, las mediciones obtenidas
de esta hormona en la sangre no correlacionan de forma positiva con los niveles
de agresión en todos los individuos.
Por tanto, surge la hipótesis
de una mayor sensibilización de los receptores para los andrógenos,
particularmente en el área preóptica del hipotálamo, el septum, la amígdala y
el núcleo dorsal del rafé en sujetos agresivos (Siegel, Roelinjg, Gregg &
Kruk, 1999; Batrinos, 2012).

Generalmente se acepta que
los andrógenos tienen efectos organizadores del cerebro y la conducta siguiendo
un patrón de masculinización, lo que desde la infancia promueve el comportamiento
de lucha a través del juego y de esta manera estimula, el desarrollo de redes neuronales
sensibles a la acción de la Testosterona que facilitan la agresión.
Aunque los niveles de Testosterona
disminuyen en la edad adulta, asociándose éste descenso con menores niveles de
agresividad, la literatura describe una mayor prevalencia en hombres que en mujeres
de trastornos de conducta asociados a comportamientos agresivos (trastorno de personalidad
antisocial, psicopatía, entre otros), por lo que la hipótesis de la agresión
asociada con la Testosterona se mantiene vigente (Moya, 2010)
Adicionalmente, la Testosterona
influiría a nivel central en la actividad de la neurotransmisión
serotoninérgica. Así, los aumentos en la Testosterona, mayor en machos que en
hembras, antes del nacimiento y durante las primeras semanas de vida extrauterina
se asocia con una menor actividad serotoninérgica en núcleos hipotalámicos como
el preóptico o el tercer núcleo intersticial del hipotálamo anterior.
Alterando los niveles hormonales a
través de inyecciones de estrógenos o andrógenos se ha podido evidenciar que la
conducta agresiva se facilita por mecanismos que involucran diferentes
receptores serotoninérgicos
Por ejemplo, en un estudio de Nelson y Chiavegatto (2011), se encontró que cuando se inducía la agresión inyectando estrógenos, la conducta disminuía si se aplicaban fármacos agonistas para el receptor 5- HT1A, mientras que cuando se producía la agresión inyectando andrógenos, la conducta se inhibía al administrar fármacos agonistas para receptores 5- HT1A-1B. Lo anterior refleja la interacción entre los andrógenos y el sistema serotoninérgico en el origen de la conducta agresiva.
Vasopresina
y Oxitocina
Por su parte, la Vasopresina
y la Oxitocina también se han relacionado con la conducta agresiva. Respecto a
la primera se ha descrito que aumentos en la actividad de este neuropéptido y
de sus receptores incrementa la conducta agresiva.
Administrando agonistas de vasopresina en regiones hipotalámicas anteriores y ventrales se observó que los roedores aumentaban la respuesta agresiva, mientras que la administración de antagonistas disminuía la respuesta.
Por otra parte, la disminución
en los niveles amigdalinos de Oxitocina se ha asociado directamente con el
incremento de conductas de hostilidad, miedo y pérdida de confianza, lo que conlleva
al aumento en la aparición de conductas agresivas.
En modelos animales se ha
encontrado que las hembras Knockout para Oxitocina mostraban mayor agresión que
hembras controles, mientras que en humanos las muestras en LCR indicaron que
los sujetos más agresivos tenían menores niveles de Oxitocina que los controles
(Ragnauth, 2005; Siever, 2008).
Estrógenos:
Así como se establece la
relación entre mayores niveles de andrógenos y la agresión, apuntando directamente
al macho por ser quien en mayor proporción libera estas sustancias, se puede sugerir
que las hembras liberan en mayor proporción estrógenos, pero ¿pueden los
estrógenos estar relacionados con la respuesta agresiva? de ser así ¿habrían
diferencias en la expresión de la agresión entre machos y hembras?
Respecto a la primera
pregunta, ha comenzado a acumularse evidencia en favor de los receptores para los
estrógenos alfa y beta en la mediación la respuesta agresiva.
Sin embargo, para comprender
la agresión en hembras y su relación con los niveles de estrógenos debe tenerse
en cuenta que los ciclos hormonales femeninos son fásicos y las variaciones en
la liberación y concentración de algunas hormonas pueden alterar significativamente
la función del hipotálamo ventromedial
(estructura sensible a la acción de los estrógenos) y de esta manera aumentar la
probabilidad de ejecutar una respuesta agresiva.
Así, se ha asociado un
aumento en los niveles de agresión en las hembras en los periodos de gestación y
lactancia, donde los estrógenos y la progesterona aumentan las concentraciones,
estableciéndose una relación positiva entre la magnitud de la agresión y los
niveles de estas hormonas.
Adicionalmente, estudios con
mujeres antes de la pubertad han demostrado que aquellas que mostraron un mayor
nivel de estrógenos manifestaban un mayor nivel de agresión y que aquellas que
recibían terapia hormonal, ya que tenían un retraso para alcanzar la pubertad,
mostraban una disminución en el umbral para iniciar la respuesta agresiva
(Ramírez, 2003).
Papel
del estrés en la agresión
Anteriormente, se mencionó
que el estrés afecta la producción de andrógenos, por lo que se asocia al
estrés con la inhibición de la agresión.
La respuesta fisiológica del
estrés está mediada por la acción del eje hipotálamo-hipofisiario-adrenal (HHA),
el cual, durante una situación desafiante para el individuo favorece la
liberación de glucocorticoides, cortisol y hormona adenocorticotropa (ACTH),
las cuales tienen un efecto directo sobre el eje hipotálamo-hipofisiario gonadal
(HHG), disminuyendo la liberación de gonadotropinas, Testosterona, hormona del crecimiento
y prolactina, así como de la respuesta agresiva.
Sin embargo, si la situación
implica un desafío temporal para el individuo, los glucocorticoides se
liberarán de forma aguda favoreciendo el inicio de la agresión, mientras que si
el desafío se prolonga, tornando crónica la liberación de glucocorticoides, la
agresión se inhibirá (Moya, 2010)
En seres humanos, los
glucocorticoides también se han asociado con la agresión; de acuerdo con
algunas investigaciones llevadas a cabo con criminales antisociales se ha
encontrado a nivel basal concentraciones bajas de cortisol y a medida que aumenta
la magnitud de la agresión aumentan también los niveles de glucocorticoides.
Por lo tanto, se ha sugerido
que una baja respuesta del sistema adrenal puede aumentar la sensibilidad hacia
estímulos estresores y de esta manera facilitar la respuesta agresiva (Moya,
2010).
Un hecho que llama la
atención al estudiar los roedores es la variabilidad en la liberación de glucocorticoides
durante el ciclo luz-oscuridad, observándose modificaciones en la expresión de
la respuesta agresiva en el curso del día. De esta manera, se ha podido
demostrar que las ratas son más agresivas durante la fase de mayor liberación
de glucocorticoides y menos agresivas durante la fase de menor liberación.
Por tanto, al establecer la
relación entre glucocorticoides y agresión debe tenerse en cuenta la mayor
cantidad de variables ya que la corticosterona o el cortisol pueden en algunos
casos facilitar la agresión y en otros casos inhibirla (Haller, Millar &
Van de Schraaf, 2000).
Estudios en los que se
seleccionaron personas con trastorno antisocial de la conducta e historial
infantil de comportamientos violentos reincidentes en la edad adulta mostraron
una menor liberación de glucocorticoides a nivel basal, hecho que ha sugerido
la posibilidad de considerar los bajos niveles de cortisol en la infancia como
un factor de riesgo para el desarrollo del comportamiento agresivo (Ramírez,
2000).
Finalmente, al estudiarse la
relación entre glucocorticoides y Testosterona, cuando un individuo muestra en conjunto
mayores niveles de Testosterona y menores niveles de glucocorticoides, tiende a
presentar agresiones de mayor magnitud, razón por la cual se ha planteado estas
variaciones como marcadores para la agresión.
Referencias bibliográficas:
Brain, P.F.,
Olivier, B., Mos,J., Benton, D., Bronstein, P.M, (1998). Multidisciplinary
studies on aggression. Swansea, University of Swansea Pres
Miczek, A., De
Almeida, M., Kravitz, A., Rissman, F. De Boer, F. & Raine, A. (2007).
Neurobiology of
escalated aggrerssion andviolence. Journal of Neuroscience. 44(44),
11803-11806.
Lesch, P. &
Merschdorf, U. (2000). Impulsivity, aggression and serotonine: A molecular
psychobiological
perspective. Behavioral Sciences and the Law, 18(5), 581-604.
Marino, D.,
Bourdelat, N., Cameron, L. & Weinshenker, D. (2005). Genetic reduction of noradrenergic
function alters social memoiry and reduces aggression in mice. Behavioral
Brain Research,
161, 197-203
Moya, L. Herrero, N. &
Bernal, C. (2010). Bases neuronales de la empatía. Revista Neurología, 50,
89-100.
Moya, L. (2010).
Psicobiología de la violencia.España: Editorial Pirámide.
Moyer, K. E.
(1976). The Psychology of Aggression. New York: Harper & Row
Purves, D. (2004).
Neuroscience. Third Edition. Sinauer Associates, Inc. U.S.A.: Publishers
Sunderland,
Massachusetts.
Raine, A.,
Buchsbaum, M., Stanley, J., Lottenberg, S., Abel, L. & Stoddard, J. (1994).
Selective reductions in prefrontal glucose metabolism in murders. Biological
Psychiatry, 36, 365-373.
Raine, A.,
Stoddard, J., Bihrle, S. & Buchsbaum, M. (1998). Prefrontal glucose
deficits in murderers lacking psychosocial deprivation. Neuropsychiatry Neuropsychology
and Behavioral Neurology, 11, 1-7.
Raine, A., Lencz, T., Bihrle, S., LaCasse, L. & Colleti, P. (2000). Reduced prefrontal grey matter volumen and reduced autonomic activity in antisocial personality disorder. Archives of General Psychiatry. 57, 19-27.
Rodriguez, M.,
Chu, R., Caron, M. & Wetsel, C. (2004). Aberrant responsesin social interaction
of dopamine transporter kcnockout mice. Behavioral Brain Research, 148,
185-158.
Sustková-Fiserová,
M., Vávrová, J. & Krsiak, M. (2009). Brain levels of GABA, glutamate and aspartate
in sociable, aggressive and timid mice: an in vivo microdialysis study. Neuroendocrinology,
30, 79-84.



Comentarios
Publicar un comentario