Neurotransmisores
Neurotransmisores
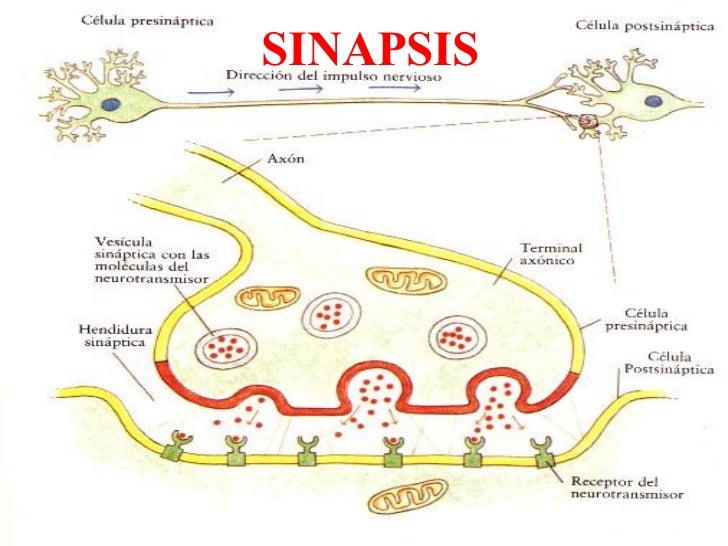
Los neurotransmisores son moléculas liberadas por las
neuronas presinápticas y son el medio de comunicación en una sinapsis química.
Se unen a receptores de neurotransmisores, que pueden
estar acoplados a un canal iónico (receptores ionotrópicos) o a un proceso de
señalización intracelular (receptores metabotrópicos).
Los neurotransmisores son específicos para el receptor al que se unen y provocan una respuesta específica en las neuronas postsinápticas, con lo que producen una señal excitatoria o inhibitoria
|
Aminoácidos GABA Glicina Glutamato Aspartato Homocisteína Taurina Péptidos opioides Metionina
encefalina Leucina
encefalina (3-Endorfina Dinorfinas Neoendorfinas
|
Aminas biógenas Catecolaminas Dopamina Noradrenalina Adrenalina Indolaminas Serotonina Histamina Melatonina Nucleótidos y nucleósidos Adenosina ATP
|



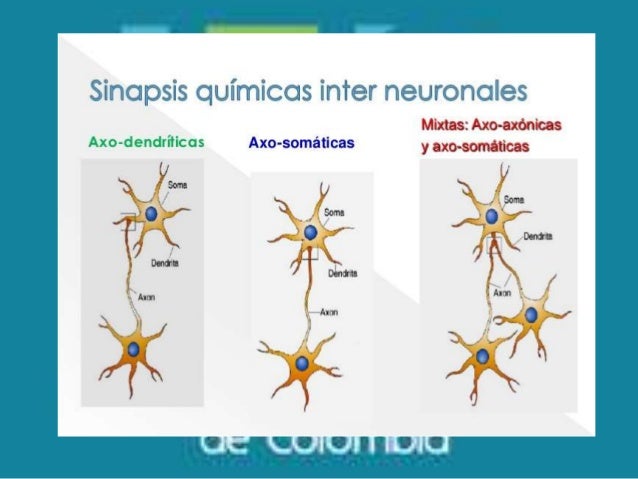
Tipos de sinapsis química del SNC
En el SNC existen diferentes tipos de sinapsis
se distinguen por la parte de la neurona que es postsináptica a la terminal
axónica.
- Si la membrana postsináptica pertenece a una dendrita, se dice que la sinapsis es axodendrítica.
- Si la membrana postsináptica pertenece al cuerpo celular o soma, la sinapsis es axosomática.
- En algunos casos la membrana postsináptica pertenece a
otro axón y estas sinapsis se denominan axoaxónicas.
- Cuando un axón presináptico entra en contacto con una espina dendrítica postsináptica, se denomina axoespinosa.
- En determinadas neuronas especializadas, las dendritas forman sinapsis unas con otras y estas sinapsis se denominan dendrodendríticas
Según su velocidad
Distintas neuronas
cerebrales liberan diferentes neurotransmisores. La velocidad de transmisión
sináptica varía considerablemente.
- Neurotransmsion rápida
En la neurotransmisión
química rápida, el receptor postsináptico es en sí mismo un canal iónico. Este
tipo de transmisión se asocia exclusivamente con los neurotransmisores de
molécula pequeña.
La unión del transmisor
estimula la apertura del canal, permitiendo un flujo de iones a través de la
membrana que altera el potencial de la misma. El proceso es rápido, ya que es directo.
Los canales iónicos en este tipo de neurotransmisión son llamados canales
iónicos modulados por ligando
o modulados por receptor; los iones implicados normalmente son sodio, potasio, calcio y cloruro
Las formas rápidas de
transmisión sináptica duran de 10-100 ms, y en la mayoría de sinapsis del SNC
son mediadas por los aminoácidos glutamato (Glu), ácido γ-aminobutírico (GABA) y la glicina (Gly).
La amina acetilcolina (ACh) media la transmisión
sináptica rápida en las uniones neuromusculares.
- Neurotransmision sináptica lenta
Las formas más lentas de
transmisión sináptica pueden durar de cientos de milisegundos a minutos; pueden
producirse en el SNC y en la periferia están mediadas por transmisores de las
tres categorías químicas.
En
la neurotransmisión química lenta, la señal es transducida por un mecanismo que
implica receptores acoplados a proteínas G. Estas proteínas y su acción se
tratarán más adelante en este capítulo. Brevemente, la unión del transmisor (frecuentemente
un neuropéptido) provoca que el receptor active una proteína G.

- Síntesis y almacenamiento de neurotransmisores
La
transmisión sináptica química requiere que los neurotransmisores sean sintetizados
y preparados para su liberación. Diferentes neurotransmisores se sintetizan de
formas diferentes.
Por
ejemplo, el glutamato y la glicina pertenecen al grupo de los 20 aminoácidos
que constituyen los componentes básicos de las proteínas ; en consecuencia,
abundan en todas las células del cuerpo, incluidas las neuronas.
En
cambio, el GABA y las aminas son producidos sólo por las neuronas que los
liberan. Estas neuronas contienen enzimas específicas que sintetizan los
neurotransmisores a partir de diferentes precursores metabólicos.
Las
enzimas productoras tanto de aminoácidos como de aminas neurotransmisoras son transportadas
a la terminal axónica, donde dirigen local y rápidamente las síntesis de transmisores
Una vez sintetizados en el citosol de la
terminal axónica, los aminoácidos y las aminas neurotransmisoras deben incorporarse
a las vesículas sinápticas.
Los transportadores se encargan de concentrar los neurotransmisores dentro de la vesícula. Estos transportadores son proteínas especiales localizadas en el interior de la membrana de la vesícula.
Liberación del neurotransmisor
La liberación del neurotransmisor se produce por la llegada de un potencial de acción a la terminal axónica. La despolarización de la membrana de la terminal hace que se abran canales de calcio dependientes de voltaje en las zonas activa

Síntesis y
almacenamiento de diferentes tipos de neurotransmisor.
a) Péptidos: En el RE se sintetiza un péptido
precursor. El péptido precursor es cortado en el aparato de Golgi,
produciéndose el neurotransmisor activo. Vesículas secretoras que contienen el péptido
emergen del aparato de Golgi.
Los gránulos de
secreción son transportados a lo largo del axón a la terminal donde se almacena
el péptido.
b) Aminas y aminoácidos neurotransmisores: Enzimas del citosol
convierten las moléculas precursoras en moléculas de
neurotransmisor. Proteínas transportadoras incorporan el neurotransmisor a las
vesículas sinápticas de la terminal, donde se almacenan.
Liberación del
neurotransmisor por exocitosis. Una vesícula sináptica rellena de
neurotransmisor en respuesta a una entrada de Ca2+ a través de canales de
calcio dependientes de voltaje libera su contenido a la hendidura sináptica por
fusión de la membrana de la vesícula con la membrana presináptica y finalmente
es reciclada por endocitosis.
El mecanismo por el que [Ca2+]i estimula la exocitosis ha sido objeto de intensa investigación. La velocidad de la liberación del neurotransmisor sugiere que las vesículas implicadas son las que están ya «ancladas» en las zonas activas.

Receptores y efectores
de los neurotransmisores
Los neurotransmisores liberados a la hendidura sináptica afectan a la neurona postsináptica uniéndose a proteínas receptoras específicas que están insertadas en la densidad postsináptica.
Aunque existen más de 100 receptores de neurotransmisor diferentes, se pueden clasificar en dos tipos: canales iónicos dependientes de transmisor y receptores asociados a proteína
Canales iónicos activados por un transmisor
Los receptores
ionotrópicos son estructuras proteicas de la membrana plasmática neuronal que
funcionana como canales iónicos específicos para determinados iones. Según sea
el tipo de ión involucrado es la naturaleza del efecto que se produce cuando
estos receptores canales se abren. Al ser canales iónicos este tipo de
receptores participan en las respuestas rápidas, excitadoras o inhibidoras, que
dan las neuronas.
Por ahora nos referiremos sólo a los receptores ionotrópicos excitadores los cuales, al permitir el paso de iones como el sodio o el calcio, producen una disminución del potencial de membrana (hipopolarización). Ello aumenta la probabilidad de generar potenciales de acción en la neurona.

Receptor- canal (por ejemplo receptor a acetil-colina)
- Espacio extracelular
- Dominio intracelular
- Poro de entrada al canal iónico del receptor
- Sitio de unión al neurotransmisor
- Canal iónico
- Estructuras formadas por aminoácidos cargados negativamente que determinan la selectividad del canal o determinados iones.
- Neurotransmisor
- Iones de sodio
Al unirse el NT al receptor puede:
~ Entrar más Na+ → genera una despolarización
(excitación)~
Entrar más Cl- → genera una hiperpolarización
(inhibición)
~ Salir más K+ → genera una hiperpolarización
(inhibición)
Receptores asociados a proteína G.
La transmisión sináptica química rápida está mediada por aminoácidos y aminas neurotransmisoras que actúan sobre canales iónicos dependientes de transmisor. Sin embargo, los tres tipos de neurotransmisor pueden actuar sobre otro tipo de receptores, los receptores asociados a proteína G, produciendo acciones postsinápticas más lentas, duraderas y mucho más variadas.
Este tipo de transmisión
implica tres pasos:
1. Las moléculas de
neurotransmisor se unen a proteínas receptoras incrustadas en la membrana
postsináptica.
2. Las proteínas
receptoras activan pequeñas proteínas, llamadas proteínas G, que tienen libertad para
moverse a lo largo de la cara intracelular de la membrana postsináptica.
3. Las proteínas G
activadas activan a su vez proteínas «efectoras».
En la parte A del esquema se muestra un receptor-canal (ionotrópico), por ejemplo al neurotransmisor glutamato. Cuando este neurotransmisor se une a su sitio de unión en el receptor, se abre el canal iónico debido a una serie de cambios conformacionales en las subunidades que lo conforman. Este cambio permite la entrada del ión sodio, lo cual provoca una respuesta inmediata en ese punto, una hipopolarización, un PEPS.
El mismo neurotransmisor también se puede unir, simultáneamente, a otro subtipo de receptor, en este caso, metabotrópico. También se va a producir acá una serie de cambios conformacionales. Como resultado de esos cambios, el receptor puede influenciar a la proteína G, ubicada en su vecindad, lo cual provoca la activación de esa proteína.
La proteína G activada puede estimular a enzimas como la adenilato
ciclasa, que induce la formación de un segundo mensajero, el adenosín
monofosfato cíclico (cAMP). Esta molécula es capaz de activar enzimas
dependientes de cAMP, como proteínas quinasas que catalizan la fosforilación de
algunas subunidades del canal iónico.
Ello provoca un cambio en la conductancia del canal, del receptor ionotrópipo,
la cual puede disminuir. La respuesta que se obtiene a través del receptor
metabotrópico demora más en aparecer y es más duradera, desde décimas de
segundo a horas.

Receptores metabotrópicos→ (más común y más lento) son receptores acopladosa proteínas G. Al unirse el NT al receptor puede:
- Abrir /cerrar canales
- Activar /inhibir segundos mensajeros
- Receptores con actividad guanilil-ciclasa intrínseca→ generan GMPc tras su activación
- Receptores con actividad tirosin-kinasa intrínsecareceptores metabotrópicos, la unión NT-receptor activa proteínas G, y éstas proteínas pueden:
- Activar/inhibir canales iónicos
- Activar/inhibir enzimas transmembrana → fosfolipasa C (PLC) o adenilatociclasas
- Activar/inhibir enzimas intracelulares
- Activar/inhibir la transcripción génica
Autorreceptores. Además de ser una parte
de la densidad postsináptica, los receptores de neurotransmisor se encuentran
frecuentemente también en la membrana de la terminal axónica presináptica. Los
receptores presinápticos sensibles al neurotransmisor liberado por la terminal
presináptica se denominan autorreceptores.
Típicamente, los autorreceptores son receptores asociados a proteína G que estimulan la formación de segundos mensajeros. Las consecuencias de activar estos receptores varían, pero un efecto común es la inhibición de la liberación del neurotransmisor y en algunos casos la síntesis del neurotransmisor..
Transducción de señales
sinápticas:
Un PA llega a la
terminación presináptica, lo que causa la abertura de los canales de Ca2+ (calcio)
regulados por voltaje.
Este influjo de Ca2+ provoca
la fusión de las vesículas llenas de neurotransmisor con la membrana y la
difusión del neurotransmisor a través de la hendidura sináptica.
El neurotransmisor se une a lo receptores postsinápticos, y los canales iónicos se abren.
El tipo de canal iónico que se abre determinará si se produce un potencial postsináptico inhibitorio (PPSI) o un potencial postsináptico excitatorio (PPSE).
Un influjo de Na+ causa un PPSE y acerca la membrana a su umbral, como se observa en la Un influjo de Cl− (cloro) causa un PPSI y aleja el potencial de membrana del umbral,
Recaptura y degradación
de los neurotransmisores
Una vez que el neurotransmisor liberado ha interactuando con los receptores postsinápticos, debe ser retirado de la hendidura sináptica para permitir que se produzca otra ronda de transmisión sináptica. Una de las maneras en que esto ocurre es por difusión simple de las moléculas del transmisor fuera de la sinapsis.
Sin embargo, en el caso de la mayoría de los aminoácidos y de las aminas neurotransmisoras, la difusión es ayudada por la recaptación del transmisor en la terminal axónica presináptica. La recaptación ocurre por la acción de unas proteínas transportadoras de neurotransmisor localizadas en la membrana presináptica.
Los 2 mecanismos de desactivacion de un neurotransmisor son la recaptacion y la inactivacion enzimatica:

Una vez en el interior del citosol de la terminal, los transmisores pueden ser destruidos enzimáticamente o bien reincorporados a las vesículas sinápticas.
Los transportadores de neurotransmisor también existen en las membranas de la glía que rodea la sinapsis, lo que ayuda a la eliminación del neurotransmisor de la hendidura.
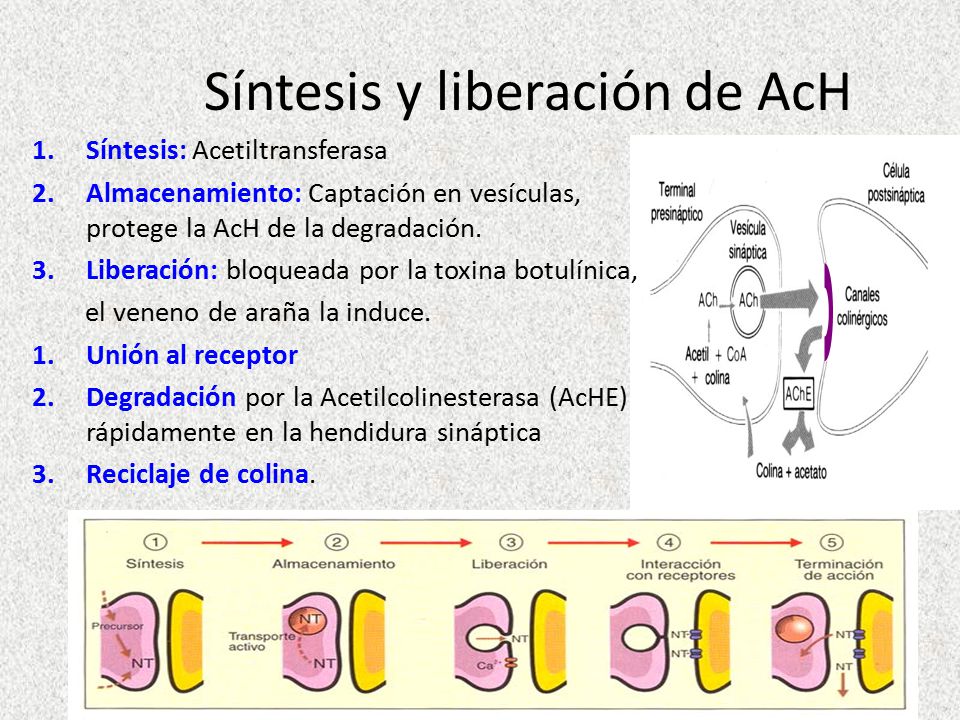
Otra manera de terminar la acción del neurotransmisor es por destrucción enzimática en la propia hendidura sináptica. Así se elimina, por ejemplo, la ACh de la unión neuromuscular. Las células musculares depositan la enzima acetilcolinesteras (AChE) en la hendidura.
La AChE rompe la molécula
de ACh, volviéndola inactiva para los
receptores de ACh.
La importancia de la
eliminación del transmisor de la hendidura no se debe subestimar.
En la unión neuromuscular, por ejemplo, una exposición ininterrumpida a altas concentraciones de ACh lleva después de unos segundos a un proceso denominado desensibilización, en el que, a pesar de la continua presencia de ACh, los canales dependientes de transmisor se cierran. Este estado desensibilizado puede persistir durante muchos segundos incluso una vez eliminado el neurotransmisor.
La rápida destrucción de
la ACh por la AChE evita normalmente que ocurra la desensibilización. Sin
embargo, si se inhibe la AChE, como ocurre con diversos gases nerviosos
utilizados como armas químicas, los receptores de ACh se desensibilizarán y la
transmisión neuromuscular fallará.
Difusión del neurotransmisor
En este proceso el NT se difunde hacia el liquido extracelular cercano aquí los astrocitos lo absorben y lo regresan a las neuronas o ser enviado al espacio extracelular al torrente sanguíneo.




Gracias!
ResponderBorrar